일주기 리듬과 건강 : 음식섭취 시간제한과 규칙적인 운동의 영향
Circadian Rhythm and Health : Influence of Food Intake Timing and Regular Exercise
Article information
Abstract
인체 항상성 유지를 위해 하루 약 24시간 동안의 주기를 갖고 변화하는 내인성 과정을 일주기 리듬이라고 한다. 일주기 리듬은 시교차상핵을 통한 내인성 중추 명령에 의해 주요하게 조절되는 것으로 알려져 있는데, 현대 사회의 현상들은 중추적 일주기 리듬을 장기적으로 교란(오정렬)시킬 수 있어 각종 질병의 유발에 기여할 수 잇는 것으로 알려지고 있다. 반면, 최근 연구들은 운동이나 식사와 같은 외인성 요인들 또한 말초에서의 일주기 리듬을 독립적으로 변화시킬 수 있다고 보고하고 있어, 중추적 일주기 리듬의 오정렬을 개선 또는 예방하는 방법으로 규칙적인 신체활동과 음식섭취 시간대 제한 등이 강조되고 있다. 따라서, 본 논문은 현재까지 연구된 일주기 리듬의 조절 기전을 살펴보고, 일주기 리듬의 오정렬과 질병과의 관계성을 정리하였다. 또한, 규칙적인 운동과 음식섭취 시간 제한을 통한 일주기 오정렬의 예방 또는 개선에 관한 연구들을 고찰해봄으로써, 건강, 신체기능 및 수명에 직접적인 영향을 미칠 수 있는 일주기 생물학 연구의 가치를 확인하였고, 일주기 리듬 유지 및 개선을 위한 규칙적인 운동과 적절한 음식섭취 습관의 과학적 근거를 소개함으로써, 일주기 리듬 유지에 어려움을 겪는 현대인들에게 규칙적인 운동과 건강한 식습관의 중요성을 강조하고자 하였다.
Trans Abstract
Circadian rhythm (CR) is an intrinsic process that changes in a cycle of approximately 24h/day to maintain body homeostasis. It is mainly controlled by the central command through the suprachiasmatic nucleus, and modern society features can disturb the central CR, contributing to various diseases. Recent studies have provided evidence that extrinsic factors, such as regular physical activity (RPA) and timerestricted feeding (TRF), can also alter the CR peripherally, emphasizing RPA and TRF as the non-therapeutic methods for circadian misalignment (CM). Therefore, this review scrutinizes the regulatory mechanism of CR and summarizes the relationships between CM and various diseases. In addition, by reviewing studies investigating the prevention or improvement of CM via RPA and TRF, the value of circadian biology research that can directly affect health, physical function, and lifespan is summarized. By introducing the scientific evidence for RPA and TRF to maintain and improve CR, we tried to emphasize the importance of regular exercise and healthy eating habits to people in the modern world who have difficulty maintaining CR.
서론
일주기 리듬(Circadian Rhythm)이란 외부환경 변화에 대응하고 적응함으로써 항상성(homeostasis)을 유지하기 위해 약 24시간 동안의 주기를 갖고 변화하는 생체 내부에서 기인하는 내인성(intrinsic) 과정을 말한다(Wolff & Esser, 2012). 시교차상핵(Suprachiasmatic Nucleus; SCN)은 시각계를 통해 전달된 빛에 관한 정보를 해석하고, 이를 토대로 말초신경계로 신호를 보내 생체 생리 시스템을 조절하는 중추적 역할을 하는 것으로 알려져 있다. 따라서, SCN은 일주기 리듬을 조절하는 마스터 중추 시계라고 불리며, 빛이 있는 낮 시간 동안에는 신체를 활성화시키고, 빛이 없는 밤 시간 동안에는 수면을 유발함으로써, 신경계의 항상성, 에너지 보존, 체온 조절, 면역 강화 등의 신체 상태를 유지시켜주는 것으로 알려져 있다(Kajimoto et al., 2018). 따라서, 일주기 리듬의 올바른 정렬(alignment)은 신체의 건강 상태를 유지하기 위해서 매우 중요하다.
선행 연구들에 따르면, 일주기 리듬의 올바른 정렬을 위해서는 하루 7~8시간의 적절한 수면이 필요하며, 부족한 수면시간(6시간 미만)은 일주기 리듬의 오정렬(circadian misalignment; CM)을 유발하는 것으로 알려지고 있다. 그 결과 암, 비만, 당뇨, 고혈압, 심혈관 질환과 같은 각종 질병들의 유발에 기여할 수 있고, 기분장애나 우울증과 같은 심리적 영향, 기억력 저하 등을 초래할 수 있는 것으로 보고되고 있다(Itani et al., 2017; Thun et al., 2021). 특히, 현대사회는 초과근무, 밤 시간대의 스마트폰 사용이나 TV시청과 같은 야간 시간대 빛에 대한 노출 증가로 인하여 SCN을 통한 일주기 리듬의 조절이 교란되어 수면 방해 또는 수면 시간의 단축을 유발함으로써 결국 잦은 CM 현상이 반복되고 있다(Walker et al., 2020). CM으로 인한 수면 부족을 개선하는 방안으로는 대표적으로 빛에 대한 노출 조절과 약물치료가 있지만, 약물치료의 경우 일시적인 효과만 나타나기 때문에 궁극적으로 CM을 해결하기에는 한계가 있다(Van Someren, 2000; Yang, 1998). CM 해결을 위한 약물치료 외 최신 연구들을 살펴보면, 규칙적인 신체활동(regular physical activity; RPA)과 특정 시간대로 제한한 시간제한섭식(time-restricted feeding; TRF)이 대표적이다. RPA와 TRF는 말초 조직에서의 일주기 리듬에 관여하는 유전자(일주기 시계 유전자, circadian clock gene; CCG)의 발현에 독립적으로 영향을 미칠 수 있음으로써, SCN을 통한 중추적 CM의 부정적 효과를 방지하거나 개선시킬 수 있다는 연구가 활발히 진행되고 있다(Hower et al., 2018). 말초 시계는 말초 조직이나 세포단위에서의 일주기 리듬을 조절하는 역할을 할 수 있고, SCN의 기능 부재 시에는 말초 시계가 독립적으로 주기를 조절할 수 있는 것으로 보고되고 있다(Gnocchi & Bruscalupi, 2017). 따라서, RPA와 적절한 식사습관을 통해 궁극적으로 CM과 연관된 당뇨, 고혈압, 심혈관 질환 등 신체에 부정적인 영향을 미치는 요인들을 개선시키는 효과가 있을 수 있는 가능성이 최근 많은 주목을 받고 있다(Hannemann et al., 2020; Longo & Panda, 2016).
하지만, 현재까지의 일주기 리듬과 건강과의 관계에 관한 연구들은 주로 SCN에서의 빛을 통한 내인성 과정에 초점으로 맞추고 있으며, 식사나 운동 같은 외인성 요인에 의한 말초에서의 일주기 리듬 재설정 메커니즘에 관한 연구는 초기 단계라 할 수 있다. 따라서 본 리뷰는 현재까지 연구된 일주기 리듬의 중추적 조절 기전과 운동, 식사와 같은 외인성 요인에 의한 일주기 리듬 변화 효과와 말초조직에서 나타나는 현상들을 살펴보고, 이를 토대로 앞으로의 일주기 리듬을 고려한 체육과학 연구의 중요성에 대해 고찰해보고자 한다.
본론
일주기 리듬
생명체는 태양의 일주(diurnal) 운동에 의한 외부의 환경 변화(빛)에 대응하여 하루 24시간 주기 내 유전자와 호르몬의 발현을 지속적으로 변화시키는 일주기 리듬을 가지고 있다(Fisk et al., 2018; Wolff & Esser, 2012). 일주기 리듬은 식물과 동물에 이르러 지구의 다양한 유기체에 존재하며 일주기 마스터 시계 또는 중추 시계라 불리는 SCN에서 담당하여 조절하는 것으로 알려져 있다(Hower et al., 2018). SCN은 인체가 일정한 주기를 가질 수 있도록, 중추적 명령을 말초에 전달함으로써 SCN과 말초조직의 리듬을 동기화 시킨다(Yoo et al., 2004). SCN의 핵 내에서는 유전자 발현 변화에 기반한 분자들의 발현 변화 리듬이 존재한다. 이러한 일주기 조절 분자 네트워크를 통해 호르몬 분비와 같은 내인성 프로세스가 조절됨으로써 체온, 수면-각성 등의 일주기 리듬을 외부환경에 맞춰 신체를 동기화한다. 이러한 외부 환경에 의한 신호를 “Zeitgeber”(독일어로 “Zeit”는 시간, “Geber”는 부여자)라고 명칭한다(Li & Zhang, 2015). 빛은 가장 강력하고 대표적인 Zeitgeber로 알려져 있지만, 포유류에는 빛 이외에도 운동, 음식 섭취와 같은 외인성 환경 요인이 말초 세포의 일주기 리듬을 조절할 수 있기 때문에, Zeitgeber로 작용할 수 있는 것으로 밝혀지고 있다(Wolff & Esser, 2012). 그러므로, 일반적으로 관찰되는 일주기 리듬은 SCN을 포함한 내재적인 중추 시계와 외인성 요인에 의해 영향을 받는 말초 시계의 끊임없는 상호작용을 통해 구현된다고 할 수 있다(Son et al., 2010).
일주기 리듬을 조절하는 분자 네트워크
포유류의 일주기 분자 네트워크는 계층의 구조로 구성 되어있고, 가장 상위 계층 기관은 SCN이다(Ko & Takahashi, 2006). 동물 연구을 통해 단백질 코딩 유전자(protein-coding genes)들의 약 43%가 말초 조직에서 일주기 리듬을 나타냈음이 밝혀졌고, 주로 새벽 또는 해질녘에 발현이 최고점 또는 최저점에 달하는 것이 보고된 바 있다(Zhang et al., 2014). SCN은 핵 내에서 전사-번역-피드백 순환 고리(transcription-translation feedback loop; TTFL) 과정을 통해 말초조직에 신호를 전달하게 된다. 포유류의 경우, 가장 상위의 전사인자(transcription factor)는 CLOCK(circadian locomotor output cycles kaput)과 BMAL1(aryl hydrocarbon receptor nuclear translocator-like protein 1)로 알려져 있으며, 이들 전사인자에 의해 조절되는 대표적인 하위 유전자로는 Period1, 2, 3(PER1, PER2, PER3), Cryptochrome1, 2(CRY1, CRY2) 및 핵 수용체 계열의 RORα(RAR-related orphan receptor alpha)와 REV-ERBα(nuclear receptor subfamily 1 group D member 1)가 있다(Lee & Kim, 2013). SCN 내 주요 분자 조절 네트워크로 기본적인 TTFL의 순환 구조는 <Figure 1>과 같다.

Molecular networks of circadian rhythm via the suprachiasmatic nucleus in response to diurnal motion
상위 단계에서 CLOCK과 BMAL1 단백질이 결합하여 이합체(dimer)(CLOCK:BMAL1)를 형성하고, 형성된 이합체는 전사인자로서 PERs 및 CRYs 하위 유전자들의 프로모터(promoter) 구역에 존재하는 enhancer box 부위에 결합하여, 하위 유전자들의 전사를 활성화시킨다. PERs 및 CRYs는 전사-번역 과정을 통하여 단백질로 생성되며, 생성된 PERs 및 CRYs 단백질들은 세포질에서 다시 이합체를 이루어 시상하부-뇌하수체-부신축(hypothalamic-pituitary-adrenal axis; HPA axis)을 활성화시켜, 신체가 활동기를 갖게 한다. 활동기를 유발한 후 이들 이합체는 세포질에서 분해 되어 다시 다량체(polymer)화 되며 서서히 축적된다. 축적된 각각의 PERs 및 CRYs 단백질은 이후 비활동 시간대에 카제인 키나제(casein kinase I isoform epsilon; CK1ε/δ)와 결합하여, 다시 SCN의 핵 내부로 이동하여 CLOCK:BMAL1 이합체의 전사 활성을 억제하기 때문에, 자신들의 유전자 발현을 조절하는 음성 피드백 순환 고리(negative feedback loop)의 역할을 하게 된다(Gallego & Virshup, 2007; Kwon et al., 2006). 세포 내 PERs 및 CRYs 단백질의 농도가 일정 역치 이하로 감소하면, 다시 CLOCK:BAML1이 활성화됨으로써, PERs 및 CRYs단백질의 농도가 증가한다. 이러한 분자 조절 네트워크를 중추적 핵심 고리(central core loop, CCL)라고 한다(Son et al., 2017). 반면, CCL의 폭주 등에 대비하는 추가 분자 네트워크가 최근 보고된 바 있는데(Lee et al., 2016; Lee & Kim, 2013), CLOCK:BMAL1 이합체에 영향을 받는 하위 유전자 RORα 및 REV-ERBα 유전자들이 추가적인 CCL 조절인자로서의 역할을 하는 것으로 보인다. CCL을 통해 생성된 RORα 및 REV-ERBα 단백질은 BMAL1 유전자의 프로모터에 위치한 REV-ERB-response element (RORE)에 전사조절 인자로 작용해, BMAL1 유전자의 발현을 조절할 수 있다. RORα은 BMAL1 유전자 발현을 활성화시키고, REV-ERBα는 BMAL1 유전자 발현을 비활성화하는 것으로 알려져 있다. 이들 분자들의 네트워크를 보조(auxiliary) 고리 또는 CCL의 주기 및 진폭을 안전하게 보조하는 역할을 담당하기 때문에 안정화(stabilizing) 고리라고 부른다(Lee et al., 2016; Lee & Kim, 2013). 추가적으로, TTFL가 안정적으로 유지되고 필요에 따라 재설정이 되려면, 다양한 방식의 전사 후(post-transcription) 또는 번역 후(post-translation) 변형이 필요하다(St. John et al., 2014). 예를 들어, 세포질 내 PERs단백질은 CK1ε/δ에 의해서 인산화 되고, CRYs단백질은 AMP-activated protein kinase(AMPK)에 의해 인산화 되는데, 인산화된 결과물들은 Skp, Cullin, F-box containing complex (E3 Ubiquitin ligase complex)에 의해 유비퀴틴화(ubiquitination)를 통한 단백질 분해가 이루어짐으로써, 세포질 내에서 PERs와 CRYs 단백질의 일정 농도가 유지된다(Jordan & Lamia, 2013; Son et al., 2017; St. John et al., 2014). 이렇듯 복잡한 분자 조절 네트워크의 여러 과정 중 어느 하나가 오작동하게 되면, TTFL가 붕괴하게 되어 인체는 일주기 리듬을 잃게 될 수 있다. 예를 들어, CK1ε/δ의 돌연변이를 유발한 실험용 쥐에서, 세포질 내의 PERs가 정상적으로 조절되지 못하여 24시간 주기가 약 20시간으로 단축되었다는 연구결과가 있다(Schroder & Esser, 2013; Sen & Hoffmann, 2020).
일주기 리듬 오정렬과 건강
신체가 원활한 일주기 리듬을 갖기 위해서는 보통 하루 7~8시간의 적절한 양의 수면이 필요하며, 충분한 수면은 중추신경계의 항상성 유지, 에너지 저장, 체온 조절, 면역력 강화 등 최적의 신체 상태를 유지시키는데 필수적이다(Kajimoto et al., 2018). 그러나, 현대 사회의 특성상 야간근무, 밤 시간대의 스마트폰사용, TV시청 등 야간에 빛을 받는 행위가 증가하였고, 더불어 신체의 노화 등으로 인한 SCN의 기능 저하가 나타남으로써 결국 수면의 방해를 유발하는 것으로 알려지고 있다. 장기간에 걸친 CM은 콜티솔(cortisol)과 멜라토닌(melatonin)과 같은 호르몬들의 분비 리듬을 교란시킴으로써, 염증장애를 유발하고 기분장애 및 포도당 항상성에 부정적인 영향을 미친다(Leliavski et al., 2015). 결과로 다양한 암 종양의 성장 또는 신진대사 장애, 비만, 심혈관 질환뿐만 아니라 우울증과 같은 심리적 인지 기능 및 정신 건강 악화와 같은 신체 악영향의 원인이 된다(Son et al., 2017; Taniguchi et al., 2009). 노인의 경우 40~70%가 수면장애로 고통받는 것으로 알려져 있는데, 이에 대한 원인으로 SCN의 기능 저하가 대두되고 있다(Van Someren, 2000).
CM에 의한 질병의 발생은 CCG 발현 변화와 밀접한 관련이 있는 것으로 알려져 있다(Lamia et al., 2008; Son et al., 2017; Taniguchi et al., 2009; Lamia et al., 2008). 다양한 암 환자들에게서 정상인에 비해 PER1, PER2의 발현이 감소되었음이 보고되었다(Son et al., 2017). 최근 연구에서, BMAL1 유전자 발현이 억제된 실험용 쥐에서 PER2의 돌연변이가 발생하였고, 종양의 성장이 가속화되는 것이 관찰되었다(Taniguchi et al., 2009). 또한, 간에서 BMAL1 유전자 발현이 비활성화 된 실험용 쥐에서 활동기 동안의 자발적인 신체활동 및 음식 섭취 타이밍의 일주기 리듬이 상실되었고, 관절병 증세가 관찰된 바 있다(Lamia et al., 2008). 비슷하게, CLOCK 유전자의 돌연변이를 유발한 실험용 쥐에서 근육과 근섬유 능력이 30%정도 감소되었고(Schroder & Esser, 2013), 골격근 미토콘드리아 수가 16% 감소되었음이 발견되었다(Pastore & Hood, 2013).
1. 일주기 리듬의 오정렬과 비만 및 제2형 당뇨병
비만은 심혈관 질환, 뇌졸중, 인슐린 저항성 및 2형 당뇨병 등 다양한 대사장애 및 질병과 관련이 있으며, 세계적으로 비만인구가 급증하면서 심각한 공중 보건 문제로 대두되고 있다(Zhou & Rui, 2013). 포유류는 하루 활동 단계 동안 에너지를 섭취하고 저장하며 수면단계 중 저장된 에너지를 이용해 균형을 맞추게 된다(Cipolla-Neto et al., 2014). 하지만, CM으로 인한 수면 부족은 비만, 당뇨병, 심혈관 질환 등 신진대사 증후군을 촉진시키는 것으로 알려지고 있다(Yoon et al., 2012). 최근 메타분석 연구에 따르면 정상적인 수면시간(7~8시간)과 비교하여 1시간의 수면 감소는 BMI지수 0.35kg 증가와 연관되어 있음이 나타났다(Cappuccio et al., 2008). 또한 멜라토닌을 분비하는 송과선(pineal gland)을 제거한 쥐에서 체중 증가가 관찰되었고, 멜라토닌을 재투여한 결과 다시 체중 감소가 나타남으로써, 멜라토닌이 체중조절에 중요한 역할을 한다는 점을 발견했다(Tan et al., 2011). 멜라토닌은 일주기 리듬에 영향을 받는 대표적인 호르몬인 점을 감안하면, 결국 CM에 의한 멜라토닌 분비 변화가 체중과 밀접한 관련이 있음을 알 수 있다. 이러한 멜라토닌의 체중조절 역할은 인슐린과도 관계가 있으며, 멜라토닌의 감소가 인슐린 저항성을 높이는 것으로 보고된 바 있다(Cipolla-Neto et al., 2014). 인슐린 또한 포유류의 포도당 대사에서 일주기 리듬을 보이는데, 아침에 분비가 활발하고, 저녁에는 감소하는 경향을 나타내며, 인슐린에 대한 감수성 또한 아침에 가장 최적이고 저녁으로 갈수록 서서히 감소하는 경향을 보인다(Mancilla et al., 2020). 반대로, 당뇨병 환자는 역전된 인슐린 분비 및 감수성 일주기 리듬을 보여 저녁에 상대적으로 좋아지지만, 야간 및 이른 아침 시간대에 악화됨으로써 이러한 일주기 리듬의 방해가 여러 합병증 유발에 기여 할 수 있다. 쥐로부터 분리한 췌장의 β세포들의 약 27%가 일주기 리듬을 나타냈으며, 이를 통한 인슐린의 분비 유도가 관찰되었다(Perelis et al., 2015). CM의 경우 인슐린 저항성 증가를 유발했으며, 포도당 항상성에 부정적인 영향을 미치고 지방 및 근육 조직에 있는 포도당 내성과 포도당 수송체 Type 4(glucose transporter type 4, GLUT4) 유전자의 발현을 감소시켰다(Froy & Miskin, 2010; Zanquetta et al., 2003; Froy & Miskin, 2010). 이외에도 CM은 SCN을 통해 분비가 조절되는 그렐린(Ghrelin), 렙틴(Leptin)과 같은 식욕 조절 호르몬 분비에도 영향을 미친다(Depner et al., 2014). 그렐린은 식욕을 증가시키는 호르몬으로 뇌하수체로부터 성장호르몬의 분비를 촉진시키고 췌장의 외분비 및 내분비기능, 탄수화물 대사, 심혈관계, 위 운동 등 생리적 기능을 조절하는 역할을 하며(Park, 2010), 반대로 렙틴은 지방 세포에 의해 생성된 식욕 억제 호르몬으로 알려져 있다(Froy & Miskin, 2010). 쥐를 활용한 선행 연구에서 CM이 그렐린을 증가시켰으며, 반대로 렙틴은 감소시켰다(Depner et al., 2014; Kettner et al., 2015; Tangestani et al., 2021; Kettner et al., 2015). 또한 건강한 성인 남녀에게서 평소의 수면-각성 주기로부터 약 4시간만의 주기 변화로도 렙틴의 수준이 10%까지 감소될 수 있는 것으로 나타났다(Nguyen & Wright, 2009). 흥미롭게도 CM은 그렐린과 렙틴의 수준을 변화시켜 비만 유도에 기여하는 것으로 알려지고 있지만, 비만이 발생한 경우에는 증가한 지방세포로부터 렙틴의 발현이 증가하여, 과도한 렙틴의 발현으로 렙틴에 대한 저항성이 증가하고 고렙틴 혈증(hyperleptinemia)이 나타나는 것으로 밝혀졌다(Dibner & Gachon, 2015; Xie et al., 2013). 이외에도 쥐를 활용한 선행연구에서 비만은 CCG중 하나인 PER1의 발현을 감소시키는 것으로 나타나 이는 내인성 프로세스를 교란시켜 궁극적으로 중추신경계의 항상성에 부정적인 영향을 미칠 수 있음이 발견되었다(Arellanes-Licea et al., 2021). 이러한 비만 예방 및 혈당 개선을 위한 일주기의 재정렬은 기본 구성 요소인 수면, 식이요법 및 신체활동을 통해 달성할 수 있다.
2. 일주기 리듬의 오정렬에 따른 호르몬 분비 변화와 기분장애 및 염증장애
CM은 PER2 및 Clock 돌연변이와 같은 CCG의 비활성화를 유발하며, 이는 우울증 및 수면부족, 또는 과다수면을 초래하여 유병률 증가에 기여할 수 있는 것으로 알려지고 있다(Melhuish Beaupre et al., 2021). SCN에 의해 분비가 조절되는 호르몬 중 하나인 콜티솔은 일주기 리듬에 있어서 매우 중요한 역할을 하지만, CM으로 인해 SCN에서의 CCG 발현이 교란되게 되면 콜티솔의 분비가 변화함에 따라 염증 장애와 우울증과 같은 기분장애를 유발하는 원인이 되기도 한다(Leliavski et al., 2015). 우울장애 환자에게서 CM은 콜티솔의 분비 패턴을 변화시키고, 멜라토닌의 감소를 유발하는 것으로 관찰되었다(Albrecht, 2013). <Figure 2>는 하루 동안 나타나는 콜티졸과 멜라토닌의 일반적인 분비 패턴과 CM에 의한 이들 호르몬의 분비 패턴 변화를 예시로 도식화 한 그림이다.
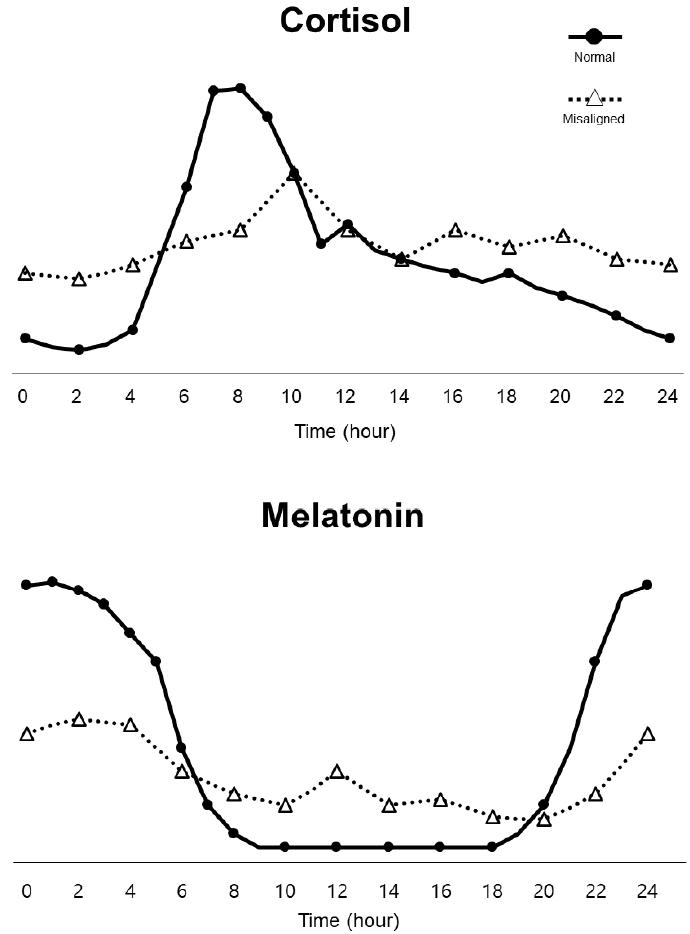
Examples of alterations in cortisol and melatonin caused by circadian misalignmentnucleus in response to diurnal motion
사전 연구에서는 우울장애 환자가 콜티솔 분비를 담당하는 HPA Axis의 높은 활성도를 보였고, 이는 아드레날린 수용체의 저항성을 증가시켜 더 높은 수준의 글루코코르티코이드(glucocorticoid)를 유도하여 궁극적으로 부신 기능의 저하에 기여함으로써 호르몬 분비 기능에 장애를 유발하였다(Holsboer, 2000). 우울장애와 더불어 양극성 장애, 계절성 장애 환자에서도 HPA axis의 비정상적인 기능으로 인해 콜티솔과 멜라토닌의 분비 패턴이 일반적이지 않은 시간대에 최고치를 보였으며 결과적으로 수면장애가 관찰되었다(Melo et al., 2016; Walker et al., 2020). CM에 의해 교란된 수면 패턴이 장기화 된 경우, 글루코코르티코이드 리듬의 변화로 인해 뇌의 해마체에서 Brain-derived neurotrophic factor(BDNF)와 그 수용체인 tropomyosin receptor kinase B의 항우울제 약물의 작용을 억제하는 모습도 보였다(Leliavski et al., 2015). 또한 수면장애로 인한 글루코코르티코이드 리듬의 변형은 항우울제 치료의 중요한 요소로 알려진 세로토닌을 포함하여 모노아민(monoamine)계열의 노르에피네프린(norepinephrine), 도파민(dopamine)과 같은 신경 전달 물질의 효율을 억제하였다(Issa et al., 2010). 또한, CM에 의한 비정상적인 호르몬 분비 패턴은 우울증 뿐만 아니라 난독증과 자폐증과 같은 질병과도 밀접한 관련이 있는 것으로 보고되고 있다. 자폐증을 가진 아동의 경우 콜티솔의 일주기 리듬에 따라 나타나는 분비, 억제의 기능이 정상적인 아동에 비해 손실이 되었음이 발견되었고, 멜라토닌의 분비 변화 또한 관찰되었다(Tordjman et al., 2014). 난독증을 가진 소아의 경우에도 아침시간대의 콜티솔 분비 저하와 콜티솔과 멜라토닌의 분비기능이 손실된 것이 관찰됐다(Huang et al., 2020). 이외에도 염증 조절에 필수적인 역할을 하는 것으로 알려진 글루코코르티코이드의 일주기 리듬이 면역반응에 중요한 역할을 한다는 점이 발견되었다(Busillo & Cidlowski, 2013). 인터류킨 6(IL6), 종양 괴사 인자(TNF-α) 및 인터류킨 1 베타(IL-1β)와 같은 염증성 사이토카인(cytokine)은 HPA axis을 자극해 글루코코르티코이드의 분비를 활성화하고, 면역 반응을 억제함으로써 염증 작용으로 인한 조직의 손상을 줄여준다. 하지만, CM은 글루코코르티코이드의 비정상적 분비를 유발하여 염증성 사이토카인을 억제 시키지 못하도록 하기 때문에 염증 장애를 유발할 수 있다고 보고된 바 있다(Wick et al., 1993). 실제로 천식, 류마티스 관절염과 같은 만성 염증 상태 질병의 환자들은 아침시간대에 글루코코르티코이드의 분비 감소가 나타났고 그에 따른 증상의 악화가 관찰되었다(Cutolo, 2019).
음식 섭취와 일주기 리듬
금식(fasting)을 포함하여 음식 섭취의 타이밍은 일주기 리듬을 조절하는 강력한신호가 될 수 있다. 음식 섭취로 유도 되는 인슐린의 분비는 간 및 지방 조직과 같은 말초 조직에 CCG의 입력 인자로 작용하는 것으로 보고된 바 있다(Bae & Androulakis, 2017). 일반적으로 식후 혈당은 저녁식사(20:00)에 비해 아침식사(08:00)시 낮게 나타나는데, 그 원인 중 하나가 인슐린과 포도당 내성의 일주기 리듬 때문이다. 인슐린은 아침에 분비가 활발하게 이루어지 때문에 아침 식사 후 혈당 수치를 낮추는데 있어서 도움을 주는 반면, 저녁시간대의 인슐린(20:00) 분비는 상대적으로 활발하지 않아, 세포로의 포도당 흡수율이 감소하기 때문에 저녁식사 후 혈당이 아침식사 후에 비해 높게 나타난다(Leung et al., 2019). 포도당 내성 또한 일반적으로 아침 시간대에 상대적으로 높은 것으로 알려져 있어 아침식사 후 상대적인 혈당 감소에 기여한다(Cipolla-Neto et al., 2014). 건강한 성인을 기준으로 식후 혈당은 아침과 비교하여 저녁 식후 12% 정도 높았고, 낮과 밤이 바뀐 12시간의 주기 이동 생활을 하는 성인(예로, 밤 근무자)에게서 전반적으로 식후 혈당이 6% 증가되었으며, 인슐린은 13%, 유리지방산의 경우에는 15%가 높게 나타났다(Morris et al., 2015). 또한 CM과 관련하여 높은 인슐린 분비로 인한 고인슐린혈증이 유발되고, 궁극적으로 인슐린 저항성을 증가시켜 비만과 제2형당뇨를 유발할 수 있다(Parameswaran & Ray, 2022). 이를 개선하는 방안으로 TRF가 주목받고 있다.
최근 연구가 활발하게 진행되고 있는 TRF는 하루 동안 전체적인 식사 시간을 특정시간대로만 제한하여 음식 섭취를 하는 것으로, 활동기 시간대, 비활동기 시간대 또는 특정 시간대를 정하는 등 다양한 방법이 있다(Hutchison et al., 2019). 사전연구들에 따르면 활동기 시간대의 TRF는 말초 조직에 호르몬 및 CCG분비시기를 변경해 일주기 리듬을 재정렬할 수 있어, CM에 의해 유발되는 비만 및 고혈당 방지에 효과적일 수 있다는 점이 발견되었다(Abe et al., 2019; Wolff & Esser, 2012). 나아가, TRF는 신체 활동량에도 영향을 미칠 수 있음이 보고된 바 있다. 실험용 쥐를 활용한 연구 결과에 따르면, 음식 섭취 시간에 따라 당뇨 그룹과 정상 그룹 모두에서 운동 및 수면의 시간대가 변경되었고(Hara et al., 2001; Oishi et al., 2004), 또 다른 연구에서는 음식 섭취 시간에 따라 쥐의 자발적인 운동 패턴이 변하였음이 보고되었다. 이 연구에서 빛을 차단된 환경에서 쥐를 관찰한 결과, 음식 제공 시간에 따라 식사 2~4시간 전에 자발적 운동이 증가함을 발견함으로써, TRF가 일주기 리듬을 조정하여 CM을 개선할 수 있다는 가설을 확인하였다(Okauchi et al., 2019). 일반적인 활동 및 수면 패턴을 지닌 건강한 대상자에게서도 TRF(08:00, 11:00, 16:00)는 활동기 시작 단계에서 콜티솔의 분비량을 증가시켜주었고, 반대로 비활동기 단계인 저녁 시간대에 콜티솔의 분비량을 감소시킴이 보고되어(Jamshed et al., 2019), 콜티솔의 일주기 리듬을 보다 안정적으로 강화시킬 수 있음을 시사하였다. 이러한 TRF 습관은 수면시간에도 영향을 미칠 수 있는 것으로 밝혀지고 있다. 음식섭취를 통해 분비되는 인슐린은 말초조직에서 일시적인 PER2의 발현을 유도했고, 이는 주기를 앞당기게 되어 빠른 비활성기를 유도함으로써 수면시간을 결과적으로 증가시켜 수면의 질 향상에 기여할 수 있다고 보고되었다(Crosby et al., 2019). 일반적인 활동 및 수면 패턴을 지닌 건강한 대상자에게서 활동기 시간대에 국한되는 10시간의 TRF는 수면 지속시간을 증가시켰으며, 설문을 통한 조사결과 수면의 만족도가 향상된 바 있다(Wilkinson et al., 2020). 반대로 비활동기 시간대의 음식(야식)을 섭취한 실험용 쥐들에게서 활동의 시작단계와 더불어 체온증가와 활동주기가 길어지고 수면의 타이밍이 지연되어 결과적으로 CM을 유발함으로써 대사질환 및 비만에 노출될 가능성을 높였다(Yoon et al., 2012). <Figure 3>은 TRF에 따른 인슐린 분비 패턴의 변화를 예시로 도식화한 그림이다.
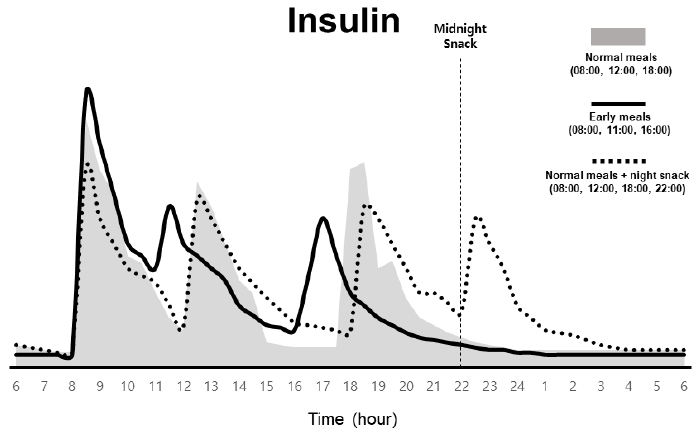
An example of insulin alterations in responses to eating time
고지방식이 및 고당식이와 같은 식습관은 비만 및 과체중을 유발하고 간, 신장, 지방 조직, 췌장과 같은 말초 조직에서 CCG와 이를 조절하는 유전자들의 분비를 감소시켜, 포도당 및 지질 대사 장애를 유발하는 것으로 알려져 있다(Honma et al., 2016; Kohsaka et al., 2007). 이러한 비만과 식습관은 자손에게도 영향을 미칠 수 있음이 동물 연구들을 통해 제시되고 있다. 고지방-고당 식이 섭취를 통해 발생한 비만한 어미쥐로부터 출산한 쥐에서 CCG의 발현 감소가 나타났으며 심장과 간에서 비정상적인 포도당 대사가 유발되었고, 궁극적으로 신진대사 장애와 비알코올성 지방간 질환의 위험이 증가하였다고 발표되었다(Borengasser et al., 2014; Mouralidarane et al., 2015; Wang et al., 2015). 반면 활동기 시간대에만 국한되는 TRF는 고지방 또는 고당 식이를 통해 나타나는 지방축적과 트라이글리세라이드(Triglyceride)의 축적을 감소시켰으며, 이는 비만 및 CCG의 변형으로부터 보호하는 역할을 할 수 있었다(Chaix et al., 2014). 이처럼 TRF는 SCN과는 독립적으로 말초 조직에서의 CCG발현 변화를 유발함으로써 CM을 개선시킬 수 있고(Kajimoto et al., 2018), 그로 인해 나타나는 신진대사에 대한 영양소의 불균형을 개선할 수 있음이 보고되고 있다(Antoni et al., 2017; Tal-Krivisky et al., 2015). 이러한 TRF의 개선 효과의 요인 중 하나는 상대적으로 긴 공복(fasting) 시간 때문이라고 할 수 있는데, 공복은 인슐린과 인슐린 성장 인자를 하향 조절함으로써, 글루카곤과 같은 내분비 호르몬을 조절하여 CM에 의해 유발되는 고혈당을 낮춰 줄 수 있기 때문이다(Jamshed et al., 2019). 최근 연구에 따르면, 활동기 시간대 TRF(08:00~17:00)를 실천한 당뇨 전 단계(prediabetes) 성인 남자 그룹에서 식후 혈당이 최대 36% 감소하였고, 인슐린 민감도 및 췌장의 β세포 기능이 회복되었음이 보고되었다(Hutchison et al., 2019). 비만한 성인의 하루 8시간 TRF는 일일 섭취 칼로리를 300칼로리 단축시켰고, 체지방 체중 등과 혈압을 감소시키는데 기여했다는 연구보고도 있어(Gabel et al., 2018), TRF는 혈당 조절 외에도 섭취 칼로리를 줄여 비만 예방에 도움이 될 수 있음이 제시된 바 있다. 또한 TRF는 비만을 통해 유발된 근육 대사 기능 및 지질 상태를 개선할 수 있는데, 비만 초파리에서 TRF가 지방축적으로 인한 미토콘드리아의 기능 장애를 개선하였고, 정상 지질 상태로 회복하고 근육 기능을 회복할 수 있다는 것이 보고된 바 있다(Villanueva et al., 2019). 또한 실험용 쥐를 활용한 사전 연구에서 TRF는 근육과 간에서 단백질 합성 지표라 할 수 있는 리보솜 단백질S6의 발현을 증가하였음을 발견함으로써 TRF가 근육 기능 또한 개선할 수 있음을 나타냈다(Chaix et al., 2014). 이처럼 활동기 시간대의 TRF는 유익한 효과를 발휘하지만, 수면시간대의 식이섭취는 TRF와 관련없이 신체에 악영향을 미치기도 한다. 선행연구에서, 비활동기(수면단계)의 식이섭취는 쥐의 간에서 CCG의 리듬을 변형시켰고, 분비 패턴을 약화시켰으며, 근육내 근위축을 유발하는 유비퀴틴 유전자 MAFbx(muscle atrophy F-box), MURF1(muscle ring finger-1) 및 Atrogin-1와 자가포식 유전자 Lc3b, Bnip3 및 GABARAPL 등의 유전자 발현을 활성화시켜 궁극적으로 근위축 유발에 기여할 수 있음이 발견되었다(Abe et al., 2019). 하지만, 현재까지의 일주기 리듬과 CM에서의 TRF 효과는 주로 동물연구로 진행되어 왔으며, 인체를 대상으로 한 연구는 매우 미흡한 실정이다(Kajimoto et al., 2018).
규칙적인 신체활동과 일주기 리듬
심혈관 질환으로 의한 사망은 아침시간대에 다소 높은 것으로 알려져 있다(Scheer et al., 2010). 나아가, CM은 하루 평균 혈압을 전반적으로 높일 수 있기 때문에, 심혈관 질환 유병률 및 질환에 의한 사망률을 증가와 밀접하게 관련되어 있다(Morris et al., 2017). 많은 선행 연구에서 밝혀졌듯이 심혈관 질환을 예방하거나 개선하는 가장 대표적인 비약물적 방법은 RPA이다. RPA는 심혈관 질환뿐 아니라 대사 질환을 포함하는 다양한 질병의 예방 및 완화 효과가 있다고 잘 알려져 있다(Lee, 2016). 최근에는 운동방법(운동 유형, 강도, 빈도 등)외에도 하루 중 어떤 시간대에 운동을 하는지에 따라 운동의 효과가 차이가 나타난다는 연구들이 많은 주목을 받고 있다. 운동 후 단시간 동안 나타나는 혈압 강하 효과(post-exercise hypotension, PEH)에 관한 한 연구에서(de Brito et al., 2015), 오전시간대 운동(07:00~11:30)과 늦은 오후~저녁시간대 운동(17:00~21:00) 모두 PEH를 유발하였지만, 오전 운동이 보다 큰 PEH를 유발하였다고 보고되었다. 이는 혈압 감소를 주 목적으로 실시하는 운동의 경우 오전 시간대가 더 효과적일 수 있다는 점을 시사한다. 그러나, 혈압은 일반적으로 아침시간대 보다 저녁시간대에 낮기 때문에, 운동시에 안전을 고려해야하는 심혈관 환자나 노약자, 임산부 등은 오전시간대 보다는(혈압강하효과는 다소 떨어질 수 있지만) 혈압이 낮은 오후~저녁시간대에 운동을 실시하는 것이 운동 중 급사(Sudden deaths)를 예방하는데 도움이 될 수 있다. 반면 제2형 당뇨환자를 대상으로 한 연구에서는 오전운동보다 오후운동이 혈당 조절에 더 큰 효과를 유도한다고 보고되었다(Heden & Kanaley, 2019). 이는 일반적인 혈당의 일주기 리듬의 경우, 저녁시간대에 식후 혈당 조절능력이 감소하고 인슐린 감수성이 악화되는 현상을 보이기 때문에, 더 취약한 저녁시간대 운동이 보다 효과적일 수 있다는 점을 시사한다. 이는 GLUT4의 변화와도 관련 지을 수 있다. 일회성 운동 후 GLUT4의 활성화는 약 24시간 동안 지속될 수 있기 때문에(Mancilla et al., 2020), 오전~낮 시간대의 운동 보다는 저녁시간대의 운동으로 혈당 조절 기능이 가장 취약한 야간시간대에 GLUT4의 활성화를 연장해 주는 것이 더 효과적일 수 있다는 것이다. 최근 연구에서는 또한 근력과 지구력 훈련을 병행하였을 경우, 오전 운동보다는 저녁 운동이 더 많은 근육량의 증가를 보이는 것으로 나타났다(Küüsmaa et al., 2016). 이는 잠재적으로 근육량이 혈당의 가장 큰 흡수원이라는 점을 감안할 때, 저녁 운동이 혈당 조절에 더 효과적이라는 점을 다시 한번 강조한다. 또한 운동은 우울증을 개선하는 것으로 알려져 있다. 장기간(1년 이상)의 우울증 환자를 대상으로 진행한 연구에서 규칙적인 운동은 우울증과 불안한 감정을 감소시켰으며 수면시간과 수면의 효율을 올려 주고, 우울증 환자가 가지고 있는 일주기 수면-각성 리듬의 불규칙성을 개선 하는데 도움을 주었다(Difrancesco et al., 2019). 또한 노인의 경우 노화로 SCN의 기능이 저하되는 것으로 보고되고 있는데, 규칙적인 운동이 노화로 인한 SCN기능저하로 유발될 수 있는 CM을 예방하는 데 도움이 될 수 있다고 제시된 바 있다(Van Someren et al., 1997).
1. 운동에 의한 수면-각성 주기 조절을 통한 일주기 오정렬의 개선
일주기 리듬은 수면 시간 및 타이밍과 밀접한 관련이 있으며, 규칙적이고 충분한 수면은 신체의 항상성 유지에 도움을 줌으로써 결과적으로 운동시 부상의 위험을 줄여주고 더 나아가 근육재생 및 복구에 중요한 역할을 할 수 있다(Chennaoui et al., 2021). 또한, 운동은 수면부족이나 노화 등으로 인해 유발되는 CM의 효과를 다소 약화시킬 수 있는 것으로 알려져 있다. 운동은 Zeitgeber로서의 역할을 하기 때문에, 이를 통한 CCG 발현 변화의 개선은 콜티솔, 멜라토닌, 체온의 변화의 일주기 리듬을 강화해 줄 수 있고 CM에 대한 치료 효과를 나타낼 수 있다(Lewis et al., 2018; Tal-Krivisky et al., 2015; Youngstedt et al., 2019). 18~40세 건강한 성인 남성을 대상으로 진행한 연구에서 수면시간을 8시간에서 4시간으로 8일간 제한한 결과 미토콘드리아의 기능저하를 유발하였고 포도당 내성이 감소하였지만, 수면을 제한하였음에도 에르고미터를 활용한 고강도 인터벌 운동(10회×60초×3세트)을 마지막 3일간 오전(10:00)에 실시한 대상자들에게서는 수면시간 부족에 의한 이러한 부정적인 현상이 감소되거나 예방되었음이 관찰된 바 있다(Saner et al., 2021). 나아가 운동이 수면시간의 부족으로 인한 미토콘드리아의 기능, 인슐린 감수성 감소, 포도당 내성 및 근형질 단백질 감소를 완화 시켜줄 수 있음이 조사된 바 있다(Morrison et al., 2022; Saner et al., 2018). 또한 실험용 쥐를 활용한 연구에서 운동이 노화로 인한 SCN의 기능저하를 예방할 수 있음이 보고되었다(Panagiotou et al., 2021). 활동기 시간대의 운동은 빛 노출을 통한 자극과 비슷하게 CCG의 분비 변화를 유발하였으며(Hower et al., 2018; Kiessling et al., 2010), 이는 결과적으로 야간시간대에 멜라토닌의 분비시기를 앞당겼고, 이로 인해 보다 빠른 수면을 유도함으로써 궁극적으로 수면 시간이 늘어났음이 보고된 바 있다. 한 연구에서는 빛이 없는 환경에서 수면-각성 일정을 하루에 20분씩 14일간 앞당긴 일반 성인에게서 활동기 시간대의 운동여부에 따른 차이를 살펴본 바가 있다. 운동(중강도, 하루당 오전 오후 각 1회)을 한 그룹에서는 체온과 멜라토닌의 분비 리듬이 약 1.6시간 단축된 반면, 그렇지 않은 집단에서는 약 0.8시간 지연되었음이 보고되었다(Miyazaki et al., 2001). 이렇듯 빛에 대한 노출이 부재한 상황에서 운동에 의해 유발되는 멜라토닌 주기의 단축은 풍족한 수면시간을 유도할 수 있어 궁극적으로 수면의 질이 향상이 되고 중추신경계의 항상성 유지에 도움을 줄 수 있다.
활동기 시간대 운동과는 반대로 야간에 실시하는 운동은 일주기 리듬의 주기를 연장 및 지연시킬 수 있다. 야간운동은 운동시에 분비되는 콜티솔의 교감신경계 활성화로 인하여 멜라토닌의 분비를 억제함으로써 주기를 지연시켰고, 이러한 현상은 밝은 조명하에 운동을 실행하였을 더 큰 지연 효과가 나타났다(Baehr et al., 2003). 한 연구에서는 적당한 조명(300Lux) 하 야간운동이 멜라토닌 분비의 시간을(55분±38)만큼 지연시켰고(Richardson et al., 2017), 또 다른 연구에서는 적당한 조명하 야간운동이 약 47분(±21.6) 정도 주기를 지연시킨 반면, 밝은 조명(5000Lux) 하 운동이 더 큰 주기 지연(56.6±15.2분)을 유발한 바 있다(Youngstedt et al., 2016). 이러한 주기의 지연은 수면부족 현상을 초래할 수 있어 (특히 밝은 조명하)은 지양되어야 하나, 반면 엘리트 선수들 같이 잦은 해외 방문으로 인한 시차 적응이 필요한 사람의 경우 야간 운동을 통하여 원활한 시차 적응을 유도할 수도 있다(Ho et al., 2005). <Figure 4>는 아침 또는 저녁시간대에 실시하는 운동에 따른 코티졸과 멜라토닌의 변화를 예시로 도식화 한 그림이다.

Examples of cortisol and melatonin changes in response to morning (06:00) or evening exercise (20:00)
운동을 통한 일주기 리듬 조절의 메커니즘으로 실험용 쥐 연구를 통해 몇 가지 제안된 바 있다. 운동은 일시적으로 PER2의 분비를 발현시키며 PER2는 단백질 CRYs와 결합하여 CRYs를 SCN내로 운반시켜주어 궁극적으로 CLOCK-BMAL1을 억제함으로써 일주기를 앞당길 수 있었다(Bruns et al., 2020). 또한 운동은 AMPK 및 HIF-1 α(Hypoxia-inducible factor 1-alpha)의 발현에도 영향을 주는 것으로 알려져 있는데(Wolff & Esser, 2019), AMPK는 CRYs, PERs 등 CCG의 분해 및 발현을 유도하였으며(Um et al., 2011), HIF-1α는 운동 시 BMAL1과 함께 발현되어 PER2의 활성화를 유도하였다(Peek et al., 2017). 또한, CRY1는 광 수용체로써 인체가 활동기에 이르렀을 때 분비되어 HIF-1α 발현을 억제하기도 한다. 메커니즘이 밝혀지지는 않았지만, HIF-1α는 PER2의 발현을 유도해 일주기를 단축할 수 있기 때문에 활성인자인 CRY1이 인체의 활동기를 오래 유지하기 위해 HIF-1α억제하는 것으로 제안된 바 있다(Dimova et al., 2019). 하지만 이러한 운동에 의해 유발되는 CCG의 변화를 통한 CM의 개선은 다소 장기간이 필요하기 때문에 일회성 운동은 CM을 개선시키기에는 부족할 수 있다. 예를 들어, 선행 연구에서 일회성 운동은 근육(슬개건)에서 CCG의 발현변화를 유발하기에는 충분하지 않았다(Yeung et al., 2019). 이에 저자들은 일회성 운동에 의해 일주기의 지연/단축이 나타나는 이유는 콜티솔과 멜라토닌과 같은 호르몬 분비의 변화와 체온이나 혈당과 같은 생리적 요인에 변화에 의해 나타나는 일시적인 변화라고 제안하였다. 운동의 강도, 시간대에 따라서 일주기 리듬의 주기 변화에 대한 분자학적 메커니즘은 아직 충분히 이해되지 않았고, 현재 진행중인 연구들이 이를 밝혀내기 위해 지속적으로 노력중이다.
2. 일주기 리듬에 따른 운동수행력 차이 및 최적의 운동 시간대
최근 운동 수행력 또한 일주기 리듬에 의해 영향을 받을 수 있다는 연구 결과들이 보고되고 있다. 많은 유형의 운동에서 아침보다는 저녁시간대에 운동 수행력이 향상된 것으로 나타났는데(Bessot et al., 2006; Fernandes et al., 2014) 이러한 연구들은 대부분 유산소 및 무산소 요소를 모두 포함하는 자전거, 달리기 및 수영과 같은 복합적인 동적 유형의 운동에서 나타났다. 반면, 저항 운동에서는 이러한 시간대별 운동 수행력 차이가 다소 덜 두드러졌고, 최대 운동 강도 이하에서만 관찰되었다(Mora-Rodríguez et al., 2012; MoraRodríguez et al., 2015).이러한 시간대별 운동수행력 차이를 설명하는 메커니즘은 아직 밝혀지지 않았지만, 예상해 볼 수 있는 요소는 몇 가지가 존재한다. 그 중 하나는 체온의 변화이다. 하루 동안 심부 체온(core body temperature)은 0.4-1.0°C 사이에서 변화를 나타난다는 것은 잘 알려져 있다(Aschoff, 1983). 하루 동안의 무산소성 파워의 변화 패턴은 심부체온의 변화 패턴과 비슷하게 나타난다고 보고되었고(Deschodt & Arsac, 2004; Kline et al., 2007), 운동 전 웜업이나 쿨다운은 하루 동안의 무산소성 파워의 변화 패턴에 영향을 주는 것으로 보고된 바 있다(Robinson et al., 2013). 따라서, 일반적인 일주기 리듬상 저녁시간대에 심부체온이 증가하는 경향을 보이기 때문에, 근육의 온도가 다소 상승되어 저녁시간대의 운동 수행능력 향상에 기여할 수 있다. 또한, 일주기 리듬에 따른 혈관의 이완과 그에 따른 혈류량에서의 차이가 하루 동안의 운동수행력 차이에 기인할 수 있다. 한 연구에서 상완의 혈관 이완 정도는 오전과 저녁시간대에 차이를 보였으며(Elherik et al., 2002) 이는 결과적으로 혈류량에 직접적인 영향을 미침으로써 일주기상 운동수행력의 차이에 기여할 수 있음을 시사한다.
또한 최근 소수의 연구들을 통해 훈련을 실시하는 시간대의 중요함이 강조되고 있는데, 주로 훈련을 실시하는 시간대에 운동 수행력이 가장 최상일 것이라는 보편적인 시각을 뒷받침하는 데이터들을 제시함으로써(Chtourou & Souissi, 2012), 경기를 앞둔 운동 선수의 경우 경기가 실시되는 시간대와 동일한 시간대에 훈련을 실시하면 도움이 될 수 있음을 시사한 바 있다. 만약, 시합이 개최되는 시간대를 알 수 없거나, 여러 날에 걸쳐 다양한 시간대에 실시하는 경우에는 아마도 아침 시간대 훈련이 더 나을 수 있다는 가능성도 제시된 바 있다(Duglan & Lamia, 2019). 이는 다소 취약할 수 있는 아침시간대의 운동수행력를 보강해 줄 수 있기 때문이다. 나아가, 식사 전 또는 후에 실시하는 운동의 차이에 대해 살펴본 연구도 존재한다. 식후 혈당은 심장질환과 관련하여 매우 중요한 위험 요소인데, 제2형 당뇨환자를 대상으로 한 연구에 따르면, 식전 운동보다 식후 운동이 식후 혈당 조절에 더 큰 이점이 있는 것으로 나타났다. 저녁 식사 이전에 실시한 운동의 경우 식후 혈당 변화를 유발하지 않았으나, 식후에 실시한 운동의 경우 식후 혈당이 감소되었으며(Colberg et al., 2009), 또 다른 연구에서는 식후 5시간 이내에 운동을 할 경우 가장 큰 식후 혈당 감소폭을 보였다는 보고가 있다(Poirier et al., 2000). 이에 대한 설명 중 하나로 골격근은 인슐린 자극에 의한 당 흡수의 50~75%를 차지하기 때문에(Frøsig & Richter, 2009), 당 흡수를 증가시키기 위한 인슐린과 근수축 사이의 시너지 효과가 나타나 식전 운동보다 식후 운동이 식후 혈당 감소에 휠씬 더 효과적일 수 있다고 제시된 바 있다(Heden & Kanaley, 2019). 혈당과 마찬가지로, 혈중지질 또한 식후 운동이 더 이를 낮추는데 더 효과적일 수 있다고 제안되었다(Gill et al., 2007). 이 외에도 일주기 리듬에 영향을 미치는 운동의 효과에 관한 분자학적, 생리학적 메커니즘은 여러가지일 수 있으나, 아직 명확한 이해가 뒷받침되지 못하기 때문에 앞으로의 관련 연구가 요구된다.
결론
현대사회의 생활 특성으로 인해 불충분한 수면의 문제점이 만연해지고 있다. 이는 만성적인 일주기리듬 변형에 노출되어 결국 CM을 유발함으로써 다양한 질병의 유병률 증가의 원인이 되고 있다. 반면, TRF는 중추적 교란으로 발생하는 CM을 개선할 수 있는 것으로 알려지고 있다. TRF는 제2형 당뇨병 환자들에게 인슐린 감수성을 향상시키고 높은 포도당 내성을 통하여 식후 혈당 및 평균적인 혈당 수치를 개선시킬 수 있고, 비만과 관련한 근기능의 저하를 예방할 수 있으며, SCN의 TTFL의 주기를 촉진시켜 수면 시간을 앞당기는데 기여할 수 있다. 이를 통해 콜티솔과 멜라토닌 같은 호르몬의 분비를 원활하게 해줌으로써 궁국적으로 수면 시간 증가 및 수면의 질 향상에 기여할 수 있다. 운동 또한 SCN으로부터 비롯되는 중추적 문제와는 별도로 독립적으로 말초조직에 CCG분비 변화를 유발하기 때문에 말초시계의 일주기 리듬을 회복시키는 역할을 할 수 있다. 식전에 비해 식후 운동은 혈당감소에 더 효과적이며, 오전의 일회성 운동이 오후에 비해 더 큰 PEH를 유발함이 보고되었다. 이외에도 정신질환을 개선시킬 수 있으며 CM을 통한 CCG리듬 변화를 통하여 항우울 효과 또한 보고되고 있다. 이처럼 TRF나 운동과 같은 외인성 요인에 의한 말초에서 일어나는 긍정적인 현상은 중추적 CM이 나타나는 사람들에게 중요한 치료목적으로 사용될 수 있을 것이다. 또한 잦불규칙적인 시차에 노출되어 SCN이 교란된 사람들에게서도 운동 또는 TRF를 통해 일주기 리듬을 보다 원활하게 회복하는데 도움을 줄 수 있고 비슷한 요인들로 인해 나타나는 신체의 변화를 인지하고 자신에 맞는 건강관리 요령을 체득해 나갈 수 있을 것이다.
Notes
CONFLICT OF INTEREST
논문 작성에 있어서 어떠한 조직으로부터 재정을 포함한 일체의 지원을 받지 않았으며 논문에 영향을 미칠 수 있는 어떠한 관계도 없었음을 밝힌다.
AUTHOR CONTRIBUTION
Conceptualization: K Lee & SK Kim; Data curation: K Lee; Formal analysis: K Lee & SK Kim; Funding acquisition: SK Kim; Methodology: K Lee & SK Kim; Projectadministration: SK Kim; Visualization: K Lee; Writing-original draft: K Lee; Writing-review&editing: SK Kim