 ISSN : 1598-2920
ISSN : 1598-2920

It has been reported that continuous exercise and bright light exposure improved resistance to stress and reduced depression and anxiety, which were attributed to enhancing neurotransmitter GABA exocytosis and stimulating neurogenesis. In this study, the effects of treadmill exercise and bright light exposure on the expression of GABAAreceptor activity, calcineurin and calcineurin calcium signaling pathway-depended NFATc4, neurogenesis-related protein Cdk5 and specific regulator factor of Cdk in neurogenesis p35 in the hippocampus of rats were investigated by western blot assay. The expression of GABAAreceptor, Cdk5 and p35 significantly increased in the exercise+light group compared to the control group and the light group. The expression of KCC2 and NFATc4 significantly increased in the exercise+light group compared to all the other groups. And the expression of calcineurin significantly increased in the exercise+light group compared to the control group. The overall results showed that exercise and bright light stimulated neurogenesis of the hippocampus in rats and had positive effects on improving the brain neuronal function.
지속적인 운동과 햇빛 치료는 스트레스에 대한 저항력을 높여주고, 우울과 불안을 감소시키며, 이러한 과정은 신경전달물질 GABA의 분비에 따라 뇌의 신경세포의 성장 및 분화 그리고 발달과정에 관여한다. 본 연구는 트레드밀 운동과 밝은 빛이 흰쥐의 해마에서 GABAA 수용체의 활성에 의해 Ca2+이 세포내 유입이 되면 calcineurin의 칼슘 의존성 경로의 활성을 통해 NFATc4 단백질 및 뇌신경세포 발달과정에 관여하는 Cdk5 단백질과 대부분의 Cdk와는 달리 신경세포 특이적 조절인자인 p35의 변화가 신경세포 생성에 미치는 영향을 알아보고자 면역얼룩법을 수행하였다. 그 결과 GABAA 수용체, Cdk5 그리고 p35는 통제집단과 빛집단에 비해, KCC2와 NFATc4는 다른 모든 집단에 비해, calcineurin은 통제집단에 비해 운동+빛집단에서 유의미하게 증가함을 알 수 있었다. 결과적으로 운동과 빛의 복합적 치료는 뇌의 해마에서 뇌신경세포 발달 과정에 관여함으로써 뇌기능을 증진시키는데 긍정적인 효과가 있음을 확인하였다.
성체 뇌에서 신경세포생성이 밝혀진 이후 신경세포의 증식(proliferation), 이동(migration), 생존(survival) 및 분화(differentiation)를 조절하는 요인들을 찾기 위한 후속 연구들이 많이 진행되었다. 흥미롭게도 트레드밀 운동은 해마(hippocampus)에서의 신경세포생성을 현저히 증가시킨다는 것으로 나타났다(Levine & Kolb, 2000; Kwon et al., 2013; Stranahan et al., 2006). 신경세포생성과 관련된 연구들에 따르면 해마에서 새롭게 형성된 신경세포들은 gamma-aminobutyric acid(GABA)라는 신경전달물질의 조절을 받아 기존의 신경세포들과 연접(synapse)을 형성하며, 자극이 풍부한 환경(enriched environment)에서 그 생존율이 현저히 증가하는 것으로 나타났다(Felice et al., 2012; Lim et al., 2015; van Praag et al., 1999)
GABA 수용체는 muscimol 등이 작용하는 A type과 baclofen 등이 작용하는 B type 수용체의 두 가지 종류가 있다. GABAA 수용체는 주로 pre-synapse에 위치하고 Cl-channel(selectivity filter)의 역할을 하며 conformational change로 channel이 열린다. 따라서, 과분극화(hyperpolarize) 시키므로, 억제(inhibitory) 역할을 한다(Sved & Sved, 1990). GABA의 주요 기능은 신경세포의 발화 억제이기 때문에 GABA는 신체를 ‘안정시키는’ 지시에서 중요한 역할을 담당한다. 따라서 GABAergic system은 과도한 자극으로부터 뇌를 보호하는 일종의 정보 필터 역할을 한다.
칼시뉴린(calcineurin)은 60KDa 촉매부위와 19KDa 칼슘 결합부위로 이루어진 이형접합 인산분해효소이다. 세포질 내 유리된 칼슘의 증가에 의해 칼시뉴린 인산분해효소는 활성화된다(Baksh & Burakoff, 2000). 칼시뉴린은 Ca2+ 증가로 활성화되고 calmodulin-binding domain을 가지고 있으며 nuclear factor of activated T cell(NFAT)를 통하여 T cell을 활성화시킨다. 인산화되어 있는 NFAT는 세포질에 존재하고 있는데 세포 내 Ca2+농도가 증가하면 칼시뉴린이 활성화되어 NFAT을 탈인산화시켜 핵으로 이동시키며 연이어 interleukin-2나 다른 임파구성장인자의 전사를 높인다.
전사인자 NFAT 단백질은 IL-2 프로모터에 결합하여 전사를 촉진하는 것이 밝혀진 이후 현재까지 5 종류가 확인되었다. 즉, NFAT family는 NFAT1 (NFATp 또는 NFATc2), NFAT2 (NFATc 또는 NFATc1), NFAT3 (NFATc4), NFAT4 (NFATx 또는 NFATc3) 및 NFAT5로 구성되어 있다. NFAT family는 복잡한 작용기전으로 면역반응에 관여하며, 특히 calcium signaling과 다른 signaling의 상호작용에 의해 면역반응을 조절한다. 또한 calcineurin/ NFAT signaling에 의해 신경영양인자 발현조절에 관여한다(Crabtree & Olson, 2002; Graef et al., 2003).
Cyclin-dependent kinase (Cdk)는 조절인자인 cyclin, 인산화효소 및 인산화효소 억제인자에 의하여 조절되며 대부분 분열중인 세포에서만 발현되어 세포주기 중에 활발히 작용하나 (Hellmich et al., 1992), Cdk5는 분열중인 세포뿐만 아니라 세포분열 후 신경세포에서도 특이적으로 발현되는 것으로 알려져 있다(Lew et al., 1994). 생화학적 특성과 염기서열은 p34cdc2 단백질 인산화효소와 유사하나, 현재까지 Cdk5의 활성을 조절하는 cyclin은 명확히 알려져 있지 않으며 신경세포 특이적으로 발현되는 p35나 p67과 결합한 경우에 활성화되고, 뇌신경세포 발달 과정에 관여하는 것으로 알려져 있다.(Lee et al., 1996; Lew et al., 1994; Tsai et al., 1994). p35는 특히 109~291 아미노산 잔기부분이 Cdk5의 활성화에 중요한 역할을 한다.(Lew et al., 1994; Poon et al., 1997)
뇌신경세포는 발달과정과 생후 초기에만 생성되며, 성인의 뇌에서는 더 이상 새로운 신경세포가 생성되지 않는다고 알고 있었지만, 1990년대 후반 성체동물에서도 새로운 신경세포가 생성된다는 사실이 증명되어졌다(Eriksson et al., 1998; Gould et al., 1999a; Gould et al., 1999b). 성체 뇌에서 신경세포가 생성된다는 후속 연구들 중에 흥미롭게도 트레드밀 운동은 해마에서의 신경세포생성을 증가하는 것으로 알려져 있다(van Praag et al., 1999; Kwon et al., 2014).
빛은 체내에 흡수되어 세포에 영양을 주고 에너지 순환을 촉진시키며, 부족으로 인해 골다공증과 같은 여러 질병을 야기 시키며(Dean, 2012), 비타민 D의 생성을 촉진시킴으로써 질병 치료에 대한 긍정적인 효과를 가진다(Yuen & Jablonski, 2010). 뿐만 아니라 빛을 통해 체내기능도 활성화되고 운동능력도 향상시킨다. 또한 인체에서 골대사와 칼슘의 항상성 유지에 있어 중요한 기능을 담당하며, 세포 증식 및 세포 분화의 조절, 면역기능 조절 등에 관여한다(Lips, 2006). 더불어 본 연구팀의 이전 연구결과를 보면 운동과 빛의 복합처지가 우울증과 관련하여 뇌의 신경전달물질인 세로토닌 분비를 더욱 더 촉진시키고, 신경생성 변화에도 영향을 줌으로써 빛이 운동과 함께 뇌의 신경 변화에 관여를 한다는 사실을 증명하였다(Kwon et al., 2013; Kwon et al., 2014a, 2014b).
따라서 뇌의 해마에서 새롭게 형성된 신경세포들은 GABA(gamma-aminobutyric acid)라는 신경전달물질의 조절을 받아 기존의 신경세포들과 시냅스를 형성하며, 시냅스 가소성을 더욱 더 용이하게 유도할 것이다.본 연구에서는 운동이나 빛과 같은 자극이 뇌의 해마에서 GABA 수용체를 활성화 함으로써 뇌 신경세포 발달에 중요한 인자인 Cdk5, p35 유전자의 발현 변화를 분석하고자 하였다.
본 연구의 실험동물은 생후 5주령 된 체중 160±10g의 수컷 Sprague-Dawley 계열의 흰쥐(KOATECH, Korea)를 사용하였다. 실험동물은 실험 전 일주일 동안 실험실 환경 적응 기간을 거치게 하였고, 깨끗한 사육케이스(28㎝×42㎝×20㎝)에 넣고 항온(22±2℃), 항습(60±10%)이 유지되며, 12시간 간격으로 낮과 밤을 교대시키는 동일한 환경으로 G대학의 청정동물실에서 사육되었으며 모든 실험 절차는 동물실험 윤리위원회의 승인을 받았다(GAR-110808-R0032). 집단분류는 무작위 표본추출에 의하여 통제집단(n=5), 운동집단(n=5), 빛집단(n=5), 그리고 운동+빛집단(n=5)으로 분류하였다. 집단별 실험동물 수는 생명 존엄성에 따른 실험동물 최소 사용이라는 위원회의 요구가 있었고 3번 이상의 반복적인 실험(in three independent experiments)을 통해 재연성을 확립하였다.
운동을 필요로 한 집단은 소형동물용 트레드밀(PARK TECH, Korea)에서 산소섭취량의 항정상태를 유지하기 위해 점증적인 부하를 이용하여 운동하도록 하였다(Takatsu et al., 2010; Tuan et al., 2008). 점증부하 운동수행은 경사도 0°에서 초기 5분간은 2m/min 속도로 부하 하다가 그 다음 5분간은 5m/min 속도로, 그리고 8m/min 속도에서 20분간, 하루 일회씩 총 30분간 달리기 운동을 일주일에 5일, 4주간 실시하였다. 4주간의 실험처치와 위와 같은 트레드밀 속도 및 경사도는 흰쥐 뇌에서 신경세포 생성 및 신경 변화에 영향을 미치는 기간과 운동 강도로 분류된다(Kim et al., 2003; Kwon et al., 2013).
본 연구를 위한 인공 광램프(Philips, Poland)는 조도 10,000 Lux까지 낼 수 있는 백광색 조명기기로 태양광 대비 91%에 근접하는 인공태양근접광원이다. 빛 치료의 목적으로는 일반적으로 10,000 Lux의 광원에 노출하므로(Gagné et al., 2007), 사용방법에 따라 인공 광램프를 실험동물로부터 80㎝의 거리에 두고 사용하였다. 하루 일회 광원 노출 시간과 실험처치 기간은 운동을 시킨 집단과 동일하며 운동+빛집단은 광원을 쬐면서 운동을 동시에 실시하였다.
실험동물은 실험 종료 48시간 후 Zoletil 50Ⓡ(10mg/kg)을 복강 내 주사를 통하여 마취시키고 단두하여 뇌 조직의 해마를 떼어내어 액체질소에 급냉동시켜 -80℃ 냉동고에 보관하였다.
보관된 조직을 lysis buffer(50 mM Tris-Cl pH 8.0, 150 mM Nacl, 1% NP-40, 0.5% Sodium deoxycholate, 0.1% SDS, 0.02% Sodium azide, proteinase inhibitor cocktail, Sigma)로 균질화 하였다. 시료를 4℃에서 12,000rpm으로 35분간 원심 분리 한 뒤, 상층액을 추출하였다. 추출된 단백질들은 BCA kit(Poerce, Rockford, IL, USA)를 이용하여 정량하였다. 정량한 단백질들은 SDS-polyacrylamine gel에서 전기영동한 후 TRANSFER(Bio-Rad, CA, USA)를 이용하여 nitrocellurose membrane (Schleicher & Shuell GmbH, Dassel, Germany)에 이동시켰다. Blot은 5% SM(skim milk) TBST로 상온에서 2시간 blocking 후 1차 항체(GABAA R1 Antibody, 1:1000, Santa Cruz, USA; KCC2 Antibody, 1:1000, Millipore, USA; Calcineurin, 1:1000, Becton Dickinson, USA; NFATc4 Antibody, 1:1000, Santa Cruz, USA; Cdk5 Antibody, 1:1000, Santa Cruz, USA; p35/25 Antibody, 1:1000, Cell Signaling, USA)를 3% SM TBST와 희석하여 4℃에서 하룻밤(overnight) 동안 반응시켰다. 세척 후, 2차 항체를 상온에서 2시간 동안 반응시켰다. Enhanced chemiluminescence(ECL)와 LAS-4000(Fujifilm, Japan)을 이용하여 발색하였다. 밴드의 크기를 동일 조건으로 비교하기 위해 α-tubulin을 통해 확인하였다.
GABAA 수용체 단백질 발현에 트레드밀 운동과 밝은 빛 노출이 뇌의 해마에서 어떠한 영향을 미치는지 확인하기 위해 Western blot 분석법을 이용하였다<Fig. 1A>.
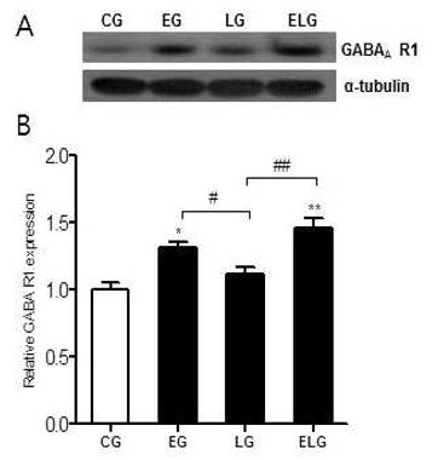
GABAA 수용체 단백질 발현의 측정 결과 통제집단 1.00±.05, 운동집단 1.31±.04, 빛집단 1.14±.05, 운동+빛집단 1.46±.07로 나타났으며, 집단 간 차이를 비교하기 위한 그래프는 <Fig. 1B>와 같다.
<Fig. 1B>에 나타난 바와 같이 각 집단 간 통계적 유의성을 검증한 결과 GABAA 수용체 단백질 발현에 유의한 차이가 있었으며(p<.01, f=12.26), 사후 검정 결과, 통제집단에 비해 운동집단과 운동+빛집단에서 유의미한 증가를 보였고 빛집단에 비해 운동집단과 운동+빛집단에서 유의미한 증가를 보였다. 그리고 운동+빛집단은 운동집단에 비해 증가한 양상은 보이나 통계적으로 유의하지는 않았다.
GABAA 수용체 활성화는 Cl-를 우선적으로 전도하는 수용체로서 K+채널을 활성화하는데 트레드밀 운동과 밝은 빛의 노출에 의해 뇌의 해마에서 KCC2 단백질의 발현 변화가 어떻게 일어나는지 알아보기 위해 Western blot 분석법을 이용하였다<Fig. 2A>.
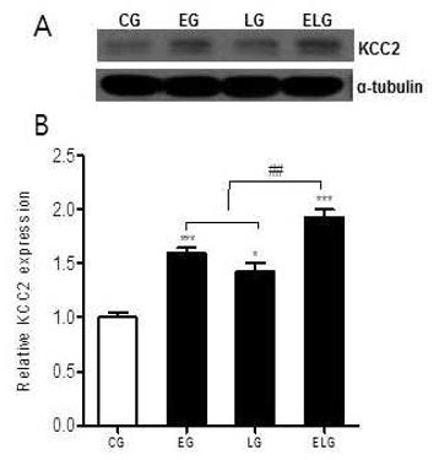
KCC2단백질 발현의 측정 결과 통제집단 1.00±.04, 운동집단 1.61±.04, 빛집단 1.43±.08, 운동+빛집단 1.93±.07로 나타났으며, 집단 간 차이를 비교하기 위한 그래프는 <Fig. 2B>와 같다.
<Fig. 2B>에 나타난 바와 같이 각 집단 간 통계적 유의성을 검증한 결과 KCC2 단백질 발현에 유의한 차이가 있었으며(p<.001, f=35.84), 사후 검정 결과, 통제집단에 비해 빛집단 그리고 운동집단, 운동+빛집단 순으로 각각의 집단에서 통계적으로 유의한 증가가 있었다. 또한, 운동+빛집단은 운동집단과 빛집단에 비해 유의한 증가가 있었다.
GABAA 수용체 활성화에 의해 Ca2+이 세포 내 유입되면 여러 가지 시그널들이 활성화되는데 그 중에서도 탈인산화 효소로 알려진 Calcineurin의 발현 변화를 알아보기 위해 Western blot 분석법을 이용하였다<Fig. 3A>.
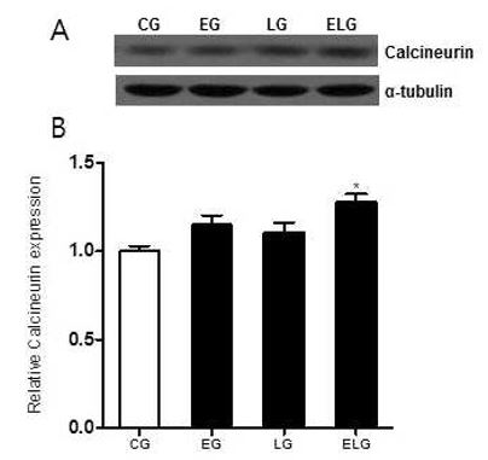
Calcineurin 단백질 발현의 측정 결과 통제집단 1.00±.04, 운동집단 1.14±.05, 빛집단 1.10±.05, 운동+빛집단 1.28±.04로 나타났으며, 집단 간 차이를 비교하기 위한 그래프는 <Fig. 3B>와 같다.
<Fig. 3B>에 나타난 바와 같이 각 집단 간 통계적 유의성을 검증한 결과 Calcineurin 단백질 발현에 유의한 차이가 있었으며(p<.05, f=5.837), 사후 검정 결과, 빛집단, 운동집단, 운동+빛집단 순으로 증가한 양상을 보였고, 운동+빛집단에서만 통계적으로 유의한 증가가 있었다.
칼슘(calcium) 시그널은 NFAT(nuclear factor of activated T cells) 단백질과 상호작용에 의해 면역반응을 조절하고 세포 분화 및 항상성을 유지하는데, 트레드밀 운동과 밝은 빛 노출에 의해 뇌의 해마에서 NFATc4단백질의 발현 변화가 어떻게 일어나는지 알아보기 위해 Western blot 분석법을 이용하였다<Fig. 4A>.
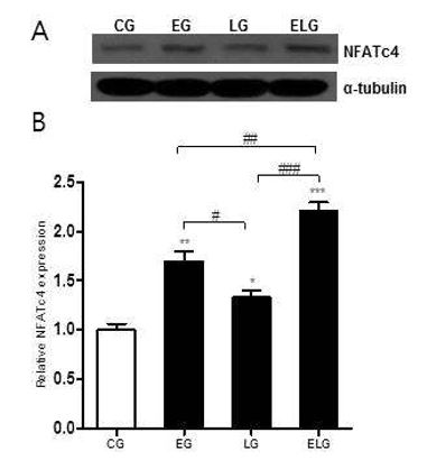
NFATc4 단백질 발현의 측정 결과 통제집단 1.00±.06, 운동집단 1.70±.01, 빛집단 1.34±.07, 운동+빛집단 2.22±.07로 나타났으며, 집단 간 차이를 비교하기 위한 그래프는 <Fig. 4B>와 같다.
<Fig. 4B>에 나타난 바와 같이 각 집단 간 통계적 유의성을 검증한 결과 NFATc4 단백질 발현에 유의한 차이가 있었으며(p<.001, f=43.34), 사후 검정 결과, 통제집단에 비해 빛집단, 운동집단, 운동+빛집단 순으로 유의한 증가를 보였고 빛집단에 비해 운동집단에서, 빛집단과 운동집단에 비해 운동+빛집단에서 유의미한 증가를 보였다.
Cdk5는 분열중인 세포뿐만 아니라 세포분열 후 신경세포에서도 특이적 발현하고 Cdk5 활성을 조절하는 p35 단백질 변화가 운동과 밝은 빛의 노출에 의해 뇌의 해마에서 어떻게 일어나는지 알아보기 위해 Western blot 분석법을 이용하였다<Fig. 5A>.
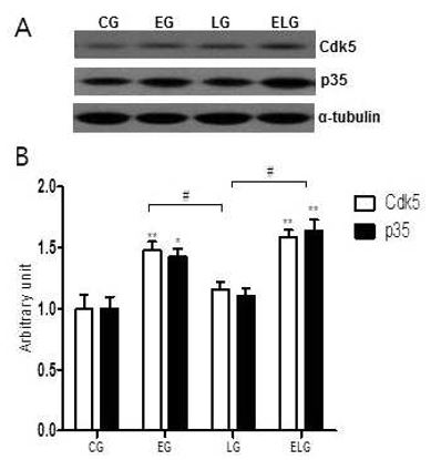
Cdk5/p35 단백질 발현의 측정 결과 통제집단 1.00±.01/1.00±.09, 운동집단 1.49±.06/1.43±.06, 빛집단 1.16±.05/1.18±.05, 운동+빛집단 1.59±.05/1.64±.09으로 나타났으며, 집단 간 차이를 비교하기 위한 그래프는 <Fig. 5B>와 같다.
<Fig. 5B>에 나타난 바와 같이 각 집단 간 통계적 유의성을 검증한 결과 Cdk5와 p35 단백질 발현에 유의한 차이가 있었으며(p<.01, f=12.23:f=13.49), 사후 검정 결과, 통제집단에 비해 운동집단, 운동+빛집단에서 통계적으로 유의한 증가를 보였고 빛집단에 비해 운동집단과 운동+빛집단에서 유의미한 증가를 보였다.
운동이 주요 억제성 신경 전달 물질 GABA를 분비하면 뇌기능 및 항우울과 항불안증의 조절에 관여 하는 것으로 알려져 있다는 기존 연구 결과들(Brambilla et al., 2003; Harvey et al., 2002; Shiah & Yatham, 1998)에 의해 트레드밀 운동과 밝은 빛이 뇌의 GABAergic 변화에 많은 관련이 있다는 가설을 제기하며 뇌의 해마에서 GABAA 수용체의 활성에 의해 세포 내 Ca2+이 유입되면 뇌 발달과정에 중요한 인자인 Cdk5와 p35 단백질 발현에 따라 뇌기능을 증진시키는 긍정적인 효과가 있을 것이라 예상하고 본 연구를 실시하였다. 게다가 빛 자극에 의해 단백질 수준이나 mRNA 수준에서 뇌 신경세포의 생성에 많은 영향을 준다는 선행 연구들(Ishida et al., 2005; Rojas et al., 2008; Shaw et al, 2010)과 마찬가지로 본 연구에서도 운동과 빛의 복합적인 처치가 뇌의 해마에서 GABAA 수용체의 활성에 의해 뇌 발달과정에 중요한 Cdk5와 p35단백질 발현을 증가시켰다. 이를 증명하기 위한 연구 결과로써 microarray analysis에서 acute한 운동 모델보다 chronic한 동물모델에서 1.5배 이상 GABAA 수용체가 증가한 분석결과가 우리의 연구 결과와 일치함을 보여주고 있다(Molteni et al., 2002). 한편 운동집단에 비해 빛집단에서 GABAA 수용체의 발현이 낮은 이유는 흰쥐 동물 모델이 야행성으로 밝은 빛에 의한 수면 방해로 수면 방추제가 감소하는데, 이는 방추제 생성에 관여하는 GABA 분비 신경세포의 감소에 의한 것으로 사료되어진다(Crowley et al., 2002). 더불어 자발적 휠 달리기는 안쪽 중격의 콜린(cholinergic)과 GABAergic 신경 세포에 의한 흰쥐 해마의 세타 리듬을 활성화 시키고, 장기간의 운동으로 유발된 동시성 뇌전도의 활성은 뇌의 화학물질을 변화시켜 신경 전달 기능에 중요한 변화를 야기 할 수 있다(Lawson & Bland, 1993). 이러한 선행연구들은 우리의 연구를 뒷받침해주고 있다.
KCC2는 세포막의 Cl-이온 기울기에 관여하는 단백질로, GABAA 수용체의 작용을 돕는다. K-Cl cotransporter인 KCC2는 세포 내 Cl-이온을 세포 밖으로 배출시킴으로써 세포 내 Cl-이온의 농도를 낮추는데, 이러한 KCC2 단백질은 연접 후 억제(postsynaptic inhibition) 및 흥분세포독성(excitotoxicity)으로 인한 신경 네트워크의 파괴를 억제하는데 중요한 역할을 한다(Tsai et al., 2011;Yu et al., 2005). 또한 신경 가소성을 조절하고 신경 손상으로 인한 KCC2의 감소는 발작을 야기하기도 한다(Kin et al., 2014; Vinay & Jean-Xavier, 2008). 이러한 연구 결과의 토대로 트레드밀 운동과 밝은 빛이 GABAA 수용체를 활성화하고, 이로 인해 KCC2 발현을 증가시킴으로써 뇌의 해마에서 신경전달물질의 조절을 받아 기존의 신경세포들과 연접을 형성하고, 자극이 풍부한 환경에서 그 생존율이 현저히 증가할 수도 있다고 유추 할 수 있으며, 신경 가소성이 더욱 더 용이하게 유도될 수 있다는 연구 결과(Schmidt-Hieber et al., 2004)와 잘 부합된다.
칼시뉴린은 인산화단백질의 세린/트레오닌잔기의 인산을 특이적으로 가수분해하는 세린/트레오닌 특이적 단백질인산가수분해효소 중 Ca2+/ calomodulin에 의해 활성조절을 받는다. 뇌에 특히 많은 점에서 단백질인산화효소와 길항하여 신경전달물질 방출의 조절, 혹은 NMDA 수용체나 이온채널의 활성조절, 또는 시냅스 가소성에 관여하고 있는 것으로 시사되고 있다(Goto et al., 1986; Matsui et al., 1987). 또한 트레드밀 운동과 같은 자극에 의해 Ca2+/ calomodulin 그리고 NFAT와 같은 신호전달기전에 의해 활성화 되면 에너지 대사와 미토콘드리아 생합성에 중심적인 역할을 한다(Akimoto et al., 2005). 이는 트레드밀 운동과 같은 자극이 미토콘드리아 기능유지와 향상에 관여하고 노화에 따른 운동 감소 능력을 예방하는 가장 기본적인 해결 방안뿐만 아니라, 흥분 전도율의 감소와 같은 신경계의 손상 등을 운동을 통해 회복할 수 있다고 간주 될 수 있다. 더불어 운동과 밝은 빛에 의해 뇌의 해마에서 Ca2+/calomodulin복합체인 CaMKⅡ 단백질의 활성이 신경발달 및 신경 보호 효과가 있다(Kwon et al., 2014b)는 것은 칼시뉴린이 통계적으로 유의미하게 증가된 본 연구의 결과를 잘 뒷받침 해주고 있다.
뇌신경세포는 발달과정과 생후초기에만 생성되며, 성인의 뇌에서는 더 이상 새로운 신경세포가 생성되지 않는다고 믿어져 왔다. 따라서 뇌신경세포는 노화가 진행됨에 따라 세포사멸에 의한 수적 감소를 보이며, 뇌 손상 시 재생이 불가능하다고 알려져 왔다. 이러한 견해는 뇌의 특정 영역에서 새로운 신경세포가 생성된다는 일부 과학자들의 발견에도 불구하고 1980년대 초반까지 지속되었다(Kaplan, 1981; Kaplan & Hinds, 1977). 그러나 1990년대 후반 새로운 실험기법들이 개발되면서 성체동물의 뇌에서도 새로운 신경세포가 생성된다는 사실이 명확히 밝혀졌다(Eriksson et al., 1998). Cdk5는 대부분의 Cdk와는 달리 신경세포 특이적 조절인자인 p35와 p67에 의해 활성화되어 신경세포에서 작용하며, 뇌신경세포 발달과정에 관여하는 것으로 알려져 있다(Lew et al., 1994; Tsai et al., 1994). 흥미롭게도 Cdk5 단백질은 달리기운동과 같은 자극에 의해 뇌의 해마에서 신경세포생성을 현저히 증가시키고 신경세포 분화과정 중 신경돌기의 성장에 필수적으로 작용할 뿐만 아니라 흰쥐 뇌 발달과정에서 신경세포 증식 및 성숙과정에 관여를 한다(Lew et al., 1994). 이는 본 연구의 운동과 빛의 복합 처리에 의해 Cdk5가 관여하고 있다는 것을 잘 나타내고 있다.
이상의 결과에서 트레드밀 운동과 밝은 빛은 GABAA 수용체의 활성에 의해 세포 내 Ca2+이 유입되면 칼시뉴린 단백질을 증가시키고 세포신호전달 NFATc4 단백질을 활성화시켜 뇌 발달과정에 중요한 인자인 Cdk5와 p35 단백질 발현에 따라 뇌기능을 증진시키는 긍정적인 효과가 있을 것이라고 생각된다.
본 연구는 5주령된 Sprague Dawley계 수컷 흰 쥐를 대상으로 4주간의 트레드밀 운동과 밝은 빛에 노출하여 면역얼룩법을 통해 해마에서 GABAA 수용체와 Cdk5 발현에 미치는 영향을 알아본 결과 다음과 같은 결론을 얻었다.
해마에서 GABAA R1, CDk5/p35 단백질 발현은 집단 간 유의한 차이가 있었으며 운동+빛집단은 통제집단과 빛집단에 비해 유의미한 증가를 보였고, KCC2, NFATc4 단백질 발현에서는 모든 집단에 비해 운동+빛집단에서 유의미한 증가를 보였다. 또한 Calcineurin 단백질 발현은 운동+빛집단에서 가장 증가한 양상은 보였으나 통제집단에 비해서만 유의미한 증가를 보였다.
따라서, 트레드밀 운동과 밝은 빛은 뇌의 해마에서 뇌신경세포 발달 과정에 관여함으로써 뇌기능을 증진시키는데 긍정적인 효과가 있을 것이라 생각된다.
Yu, S. J., Kim, J. R., Lee, C. K., Han, J. E., Lee, J. H., Kim, H. S., Hong, J. H., & Kang, S. G. (2005). Gastrodia elata blume and an active component, p-hydroxybenzyl alcohol reduce focal ischemic brain injury through antioxidant related gene expressions. Biological & Pharmaceutical Bulletin, 28(6), 1016- 1020.